O corpo humano pode ser, sem exagero, classificado entre os sistemas biológicos mais complexos do planeta. Nosso corpo consiste em bilhões de células, muitos órgãos e sistemas. Mas todo esse esplendor e diversidade surgiram devido à fusão de apenas duas células - um espermatozóide e um óvulo. Acho que não há necessidade de explicar como ocorre a fertilização e o que é necessário para isso (dica - cegonha e repolho não têm nada a ver com isso). Mas aqui estão alguns aspectos da vida do esperma por muitos anos que permaneceram obscuros. Cientistas da Universidade de Bristol, usando técnicas modernas de microscopia tridimensional, foram capazes de ver os movimentos dos espermatozoides de uma forma que antes não era possível. Como e por que meios o esperma se move, aprendemos com o relato de cientistas. Ir.
Base de pesquisa
Apesar de os espermatozoides estarem envolvidos na criação da vida muito antes do advento do método científico, sua trajetória na literatura científica começou bem recentemente - em 1677. O estudante de medicina Johann Gam compartilhou suas observações com o colega e amigo Anthony van Leeuwenhoek (1632 - 1723), que, por sua vez, examinou e descreveu em detalhes os "animais seminais" (como chamava os espermatozóides).
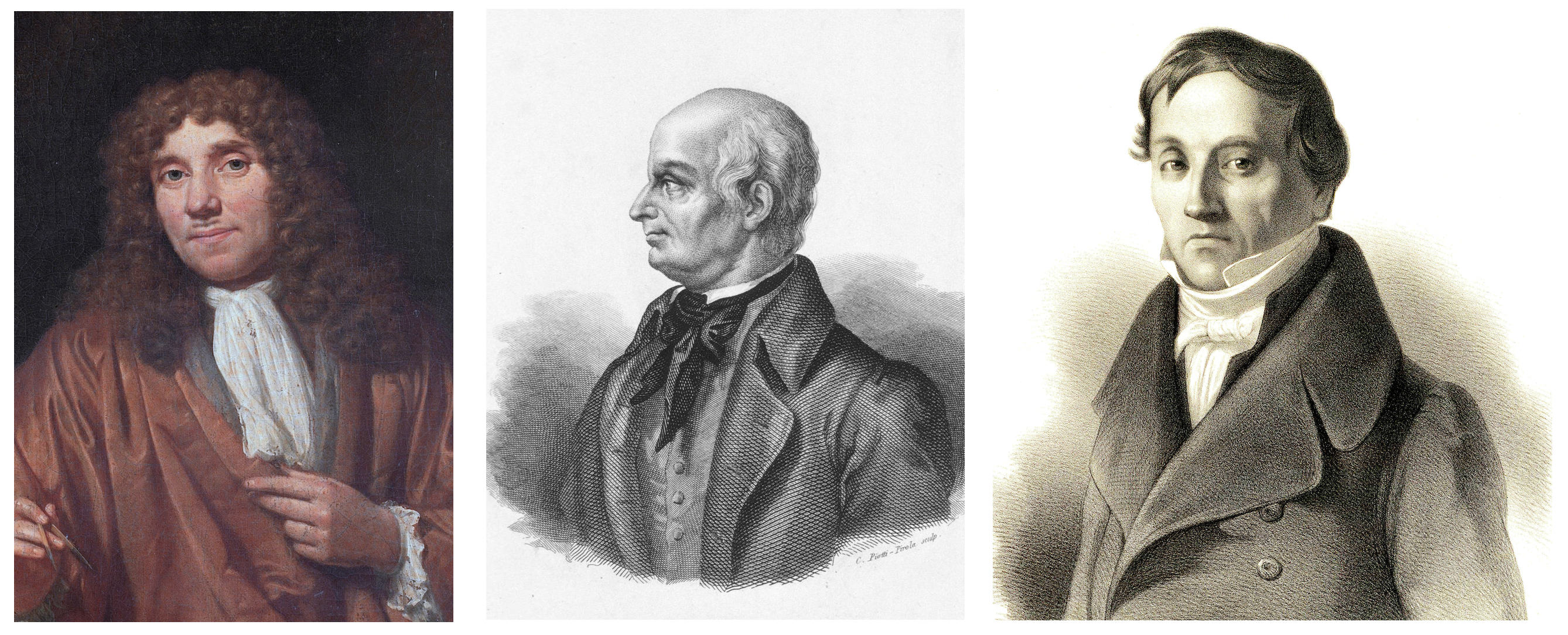
Anthony van Leeuwenhoek / Lazzaro Spallanzani / Carl Ernst von Baer
Leeuwenhoek sugeriram que essas células incomuns estão envolvidas na fertilização, mas sua teoria, embora verdadeira, foi rejeitada pela comunidade científica. Por muito tempo, acreditou-se que os espermatozoides eram parasitas e apenas o fluido seminal estava envolvido na fertilização.
Quase cem anos depois, em 1786, o fato da participação dos espermatozoides na fecundação foi comprovado por Lazzaro Spallanzani (1729-1799) em sua obra Experiencias Para Servir a La Historia de La Generación De Animales y Plantas. No entanto, suas explicações para o processo em si eram bastante vagas: ele acreditava que o óvulo já era o início de um novo organismo e que o esperma só era necessário para ativar o processo de crescimento.
O mesmo termo "esperma" foi introduzido no início do século 19 por Karl Ernst von Baer (1792 - 1876).
Independentemente do que os cientistas pensaram há vários séculos, os espermatozoides têm uma função muito clara, cuja implementação é fornecida por várias ferramentas especializadas. A principal tarefa do espermatozóide humano é passar pelo trato genital feminino, encontrar o óvulo e transferir o material genético masculino para ela.

A estrutura do espermatozóide A
célula reprodutiva masculina não pode se orgulhar de dimensões, porque é a menor do corpo humano (excluindo a cauda): as dimensões da cabeça são 5,0x3,5x2,5 mícrons (comprimento x largura x altura), o comprimento da parte do meio é 4,5 mícrons, e comprimento da cauda - 45 mícrons.
Ao mesmo tempo, o tamanho pequeno não é uma desvantagem, mas um aspecto importante para aumentar sua velocidade. No processo de maturação do esperma, seu núcleo (carrega um único conjunto de cromossomos) torna-se mais denso, a maior parte do citoplasma é descartada e apenas as organelas mais importantes permanecem na célula.
O flagelo pode ser chamado de segundo elemento mais importante do esperma depois do núcleo, ou seja, sua cauda. Pois é graças a ele que se realiza o movimento dessa célula ao longo do trato genital de uma mulher. Também é engraçado que o ambiente vaginal seja extremamente destrutivo para as células germinativas masculinas, mas o sêmen reduz parcialmente o efeito negativo no esperma. O nível de pH dentro do trato genital feminino permite que os espermatozoides se movam em direção ao útero, onde um ambiente muito mais favorável os aguarda.
Anteriormente, acreditava-se que o esperma se move para frente devido ao movimento simétrico de seu flagelo de um lado para o outro.
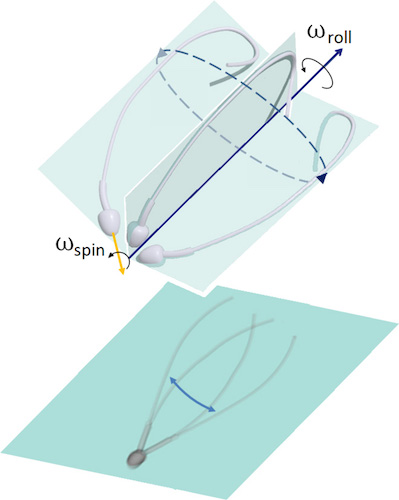
Imagem # 1: assimetria do movimento flagelar em 3D (topo); uma projeção plana do movimento do flagelo, criando uma ilusão de ótica de simetria bilateral em microscopia 2D (parte inferior).
Esta declaração foi expressa até mesmo nos dias de Levenguk. Também levou a uma idealização simétrica da forma de onda em três dimensões, muitas vezes percebida como uma espiral cônica, semelhante a um saca-rolhas em expansão.
Devido à microscopia bidimensional, muitas observações foram interpretadas incorretamente e, às vezes, completamente erradas. As afirmações sobre a simetria dos retalhos do flagelo contradizem muitas observações que mostram a assimetria estrutural dentro da estrutura do próprio flagelo.
Se o bater do flagelo do esperma durante o movimento ainda é assimétrico, então como a simetria do movimento do flagelo de um lado para o outro e o movimento da célula na direção para frente são alcançados? É esta questão a principal neste estudo.
Para obter uma resposta, os cientistas compararam observações moleculares e microscópicas que mostraram que o esperma humano usa o controle assimétrico e anisotrópico para regular o flapping flagelar. Em outras palavras, a simetria é realizada devido à assimetria: o efeito de um “topo de precessão” surge quando a cabeça gira (“fluido de perfuração”, palavras dos autores) simultaneamente e independentemente de como o flagelo do esperma gira em torno do eixo de movimento.
Resultados da pesquisa
O rápido movimento dos flagelos do esperma humano foi registrado com alta resolução espaço-temporal em 3D. Dois grupos de espermatozóides flutuantes em um fluido de baixa viscosidade foram examinados: espermatozóides que flutuam ao lado da lamela (fica no topo da amostra durante a microscopia) e longe dela.
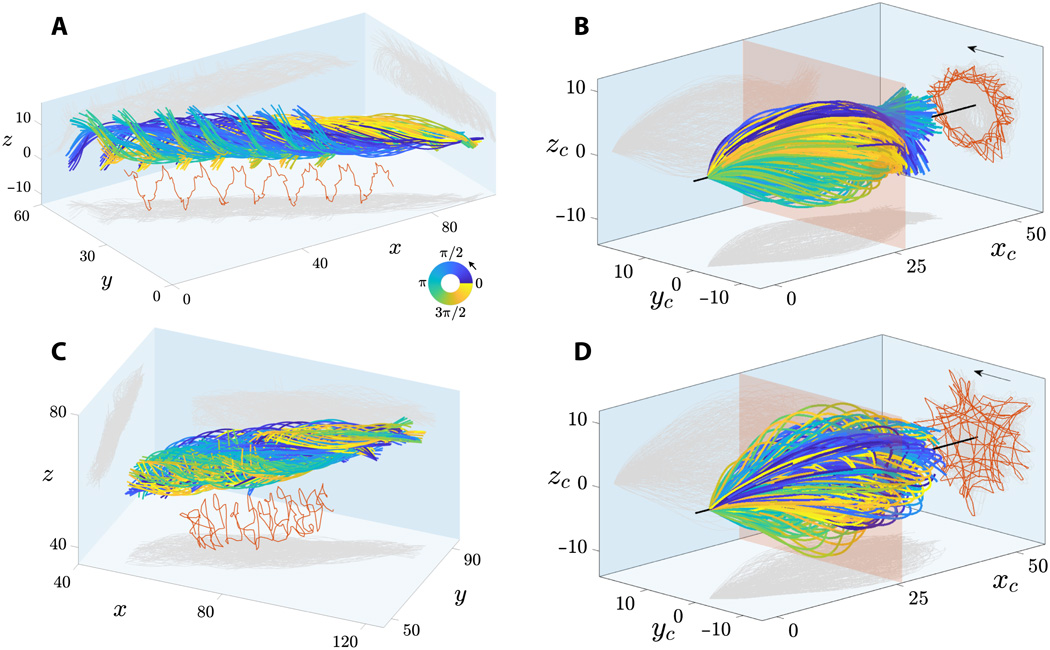
Imagem №2
Em 2A e 2C mostram flagelos de esperma flutuando perto e longe da lamela. Os retalhos do flagelo são caracterizados por um movimento de rolamento característico em torno da direção do movimento do esperma.
O movimento combinado de rotação e translação do flagelo do esperma leva a trajetórias espirais do ponto médio do flagelo com quiralidade preservada (marcado em vermelho em 2Ae 2C ). Os espermatozoides mostram rotação bidirecional em torno de seu eixo: todas as células flutuantes livremente (28 peças) giradas no sentido anti-horário quando vistas de frente (setas em 2B e 2D ) e apenas 2 células giradas no sentido horário (elas não se movem para frente devido a obstáculos no caminho deles).
Microscopia de espermatozóides flutuando ao lado da lamela (corresponde a 2A ).
Todos os espermatozóides (30 células) se moveram de forma semelhante a um topo rotativo em precessão, no qual a rotação da cabeça em torno do eixo longitudinal do espermatozóide ( rotação ω ) ocorre simultaneamente e independentemente de como o flagelo gira em relação ao eixo de movimento ( rolo ω ).
Em 2B e 2D mostra os termos do flagelo oscilante do sistema de coordenadas concomitante * , ou seja, um ponto de visualização, se move junto com o esperma, mas não o movimento rotativo em torno de seu eixo. Isso mostra que os golpes flagelares são extremamente simétricos tanto no plano ( xy ) quanto fora do plano (z ) direções, que correspondem às observações em 3D.
O quadro de referência * que o acompanha é um quadro de referência associado ao corpo em questão em um determinado momento. O corpo dentro desse sistema está imóvel. Por exemplo, um elevador em queda livre é um quadro de referência que o acompanha para um corpo que cai livremente nele, mas a Terra não é esse sistema em relação ao corpo em um elevador.A projeção da trajetória do meio do flagelo (linhas vermelhas em 2B e 2D ) mostra uma incrível matriz de padrões geométricos, de estrelas girando a triângulos, quadrados e padrões de loop com simetria polar. O padrão flagelar irregular mostrado em 2D para espermatozóides flutuando para fora da lamela também é observado para espermatozoides flutuando próximo à lamínula. Essa variabilidade nos padrões pode ser causada por uma incompatibilidade no atraso de fase entre os componentes no plano e fora do plano do flapping para cada célula, o que aumenta na linha do tempo do movimento de rolamento. Consequentemente, os padrões flagelares não são uma característica distintiva das células flutuando perto e longe da lamela.
Microscopia e modelagem do flagelo mostrando a rotação da cabeça e do flagelo em torno do eixo de movimento.
A amplitude da onda 3D é caracterizada por uma curva de envelope simétrica em forma de bala, em oposição à espiral cônica (semelhante a um saca-rolhas em expansão), que é frequentemente descrita na literatura. Os espermatozóides que flutuam para longe da lamela têm uma forma de onda mais simétrica do que as células flutuando ao lado do vidro. Assim, a lamínula adjacente é uma fonte fraca de assimetria para o flapping flagelar.
Os espermatozóides flutuando ao lado da lamela tinham um ângulo de ataque preservado de * -7 °, com uma orientação média do flagelo do esperma direcionado para a lamínula.
* — ( ) , .

Imagem nº 3
A imagem acima mostra uma comparação de golpes flagelares no quadro complementar (linha superior) e no quadro giratório que o acompanha (linha inferior).
A Figura 3E mostra a verdadeira natureza da batida flagelar vista da perspectiva do esperma, sem qualquer movimento de flutuação ou rolamento.
O quadro de referência rotativo ( 3E ) que acompanha mostra que as abas flagelares são anisotrópicas, ou seja, as características de onda em cada direção transversal (perpendicular ao plano oscilante), rotuladas de “ plano b ” (plano azul) e “ plano z ” (plano vermelho), são marcadamente diferentes.
Se compararmos as áreas cinza claro nos planos azul e vermelho em 3E, então pode ser visto que o flapping é fortemente assimétrico no plano b e é caracterizado por uma simetria esquerda-direita quebrada, lembrando um formato de C.
Esta observação está em forte contraste com os padrões simétricos observados no quadro de referência comovente em 3A .
Forma de onda flagelar em relação a um referencial fixo de laboratório (x, y, z).
A análise do componente principal (PCA) de uma forma de onda tridimensional permitiu aos cientistas decompor a onda flagelar em vários "modos de forma básicos", aqui referidos como modos PCA. A forma de onda 3D pode ser reconstruída com boa precisão com apenas dois modos de forma de onda, como visto com 3B e 3F .
Em 3C, pode ser visto que os dois primeiros modos de PCA são idênticos em forma, até uma rotação de 90 °, capturando assim a forma de espiral aerodinâmica causada pela rolagem de esperma.
No 3G , no entanto, a forma C assimétrica interna é totalmente detectada apenas pelo primeiro modo PCA. Segundo modo PCA em 3G(destacado em laranja) introduz pequenos desvios perpendiculares ao primeiro modo PCA (destacado em azul), indicando que a forma de onda pode ser decomposta em duas direções de ondulação transversais independentes que são anisotrópicas por natureza.
A análise de Fourier das varreduras permitiu reconstruir o movimento do flagelo usando apenas dois modos de Fourier em 3D e 3H . Em suma, cada sinal de varredura comum pode ser aproximado por uma soma simples de duas funções:
f r (s, t) ≈ f 0 (s) + | f 1 (s) | sin (ωt + φ (s))A primeira função f 0 (s) não depende do tempo, é chamada de "modo estático" e corrige a assimetria média do sinal ao longo do (s) arco (s) do flagelo.
A segunda função é uma onda viajante senoidal denominada "modo dinâmico", oscilando na frequência capturada pelo primeiro pico do espectro de potência do sinal.
A amplitude e a modulação de fase de uma onda viajante ao longo do flagelo são iguais, respectivamente, | f 1 (s) | e φ (s) = arg (f 1 (s)).
Assim, a fase carrega informações sobre as características da onda viajante. Por exemplo, se a fase φ (s) não muda ao longo de s, então o sinal não é uma onda viajante, mas sim uma onda estacionária.
Portanto, o modo estático captura qualquer incompatibilidade de forma de onda. Nesse caso, a linha reta preta em 3D reflete a simetria da varredura nos planos xy e xz do quadro móvel. Em contraste, no referencial rotativo ( 3H ) que o acompanha , o modo estático (curva preta) é caracterizado por uma grande amplitude assimétrica, que lembra um C. invertido.
Forma de onda do flagelo em relação ao quadro de referência que o acompanha.
Forma de onda do flagelo em relação ao quadro de referência rotativo que o acompanha.
O modo dinâmico no quadro complementar (curvas vermelhas) em 3D tem grande amplitude e é altamente simétrico nos planos xy e xz devido ao rolamento do esperma.
Mas o modo dinâmico no referencial de rolagem que acompanha (curvas vermelhas) em 3H tem uma amplitude de forma de onda reduzida e uma direção preferencial de deslocamento.
A reconstrução de Fourier da forma de onda foi realizada somando os modos estático e dinâmico (gráficos no centro em 3D e 3H ), que está em boa concordância com as observações originais (gráficos à direita em 3De 3H ).
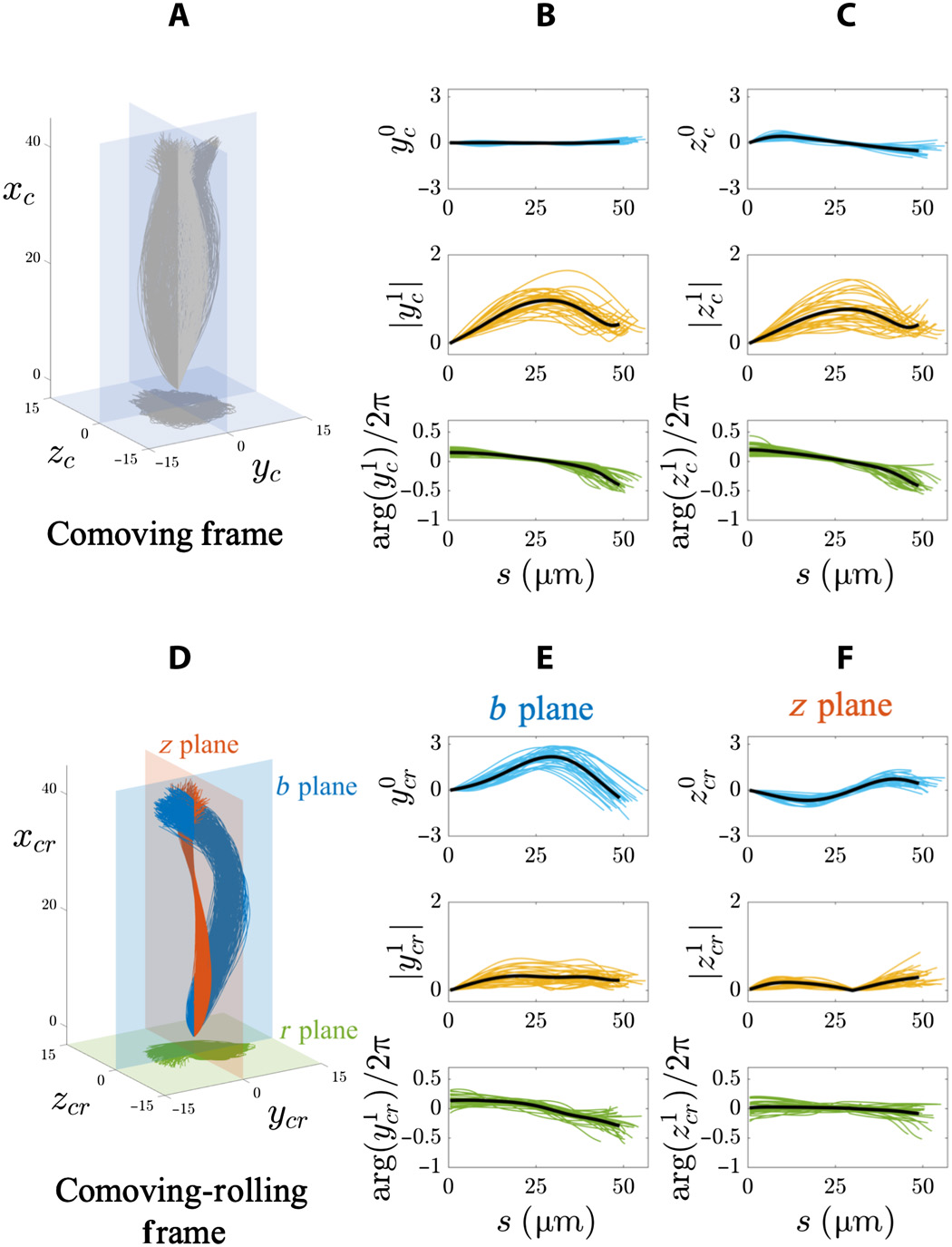
Imagem # 4 Os
gráficos acima mostram os resultados de uma análise de Fourier de um retalho flagelar 3D em uma população de espermatozoides flutuantes (20 células ao lado da lamela e 8 células de distância do vidro). No quadro de referência que o acompanha ( 4A - 4C ), as amplitudes do modo estático em ambas as direções (y c e z c ) são muito pequenas (linha superior dos gráficos em 4B e 4C ) devido à simetria das varreduras.
Além disso, a amplitude (linha do meio) e a fase (linha de baixo) dos modos dinâmicos (y c e z c ) em 4B e 4Cfixa a simetria transversal e excede a isotropia neste quadro de referência devido ao fato de que o espermatozóide gira em torno do eixo de movimento.
As características da onda viajante para ambas as coordenadas (y c e z c ) são as mesmas para todas as células de flutuação livre: frequência 4 Hz, comprimento de onda 100 μm e velocidade da onda 400 μm / s.
No sistema de referência rolante que acompanha ( 4D - 4F ), os regimes estáticos distintas nos b e z aviões mostram que as oscilações têm ainda anisotropia numa população de células de flutuação livre ( 4E e 4F ). Plano b do
modo estático (y cr) é fortemente assimétrico e inclinado para valores positivos (linha superior dos gráficos por 4E ). Mas para o plano z (z cr ), ele oscila simetricamente ao longo do comprimento do arco de forma senoidal (linha superior em 4F ).
A amplitude do regime dinâmico no b plano (y 1 cr ) aumenta até atingir um plateau (linha do meio na 4E ), enquanto o regime dinâmico na z plano (z 1 cr ) é não monótona ao longo do comprimento do arco (linha do meio na 4F ). Características da onda viajante y cr ( bplano) foram: frequência 8 Hz, comprimento de onda 145 μm e velocidade da onda 1120 μm / s. Características de uma onda viajante z cr ( plano z ): frequência 6 Hz, comprimento de onda 1526 μm e velocidade 5174 μm / s.
Pequenas mudanças de fase em grandes distâncias ao longo de todo o comprimento do flagelo requerem velocidades de propagação de ondas muito altas. Conseqüentemente, as oscilações do plano z realmente se comportam como uma onda estacionária pulsando no tempo.
Microscopia de espermatozóides não propulsores e rolantes.
A totalidade dos dados acima sugere que o flapping dos flagelos usa dois controladores transversais anisotrópicos coativos, que não são muito diferentes das ondas eletromagnéticas viajando. No entanto, cada onda de cisalhamento (y cr , z cr ) é a soma dos modos estático e dinâmico: uma onda progressiva assimétrica ao longo do plano b (azul em 4D ) e uma onda estacionária simétrica no plano z (vermelho em 4D ).
É curioso (mas não surpreendente) que a presença de uma tampa de vidro perto da trajetória da célula espermática também tenha seu efeito na amplitude de propagação das ondas.

Imagem nº 5 O
vidro reduz a amplitude de propagação das ondas devido às interações hidrodinâmicas entre o flagelo e a superfície dura da tampa de vidro.
No quadro complementar ( 5A e 5B ), a amplitude de ambos os modos dinâmicos (y c , z c ) diminui em direção ao final do flagelo do esperma perto da lamela (gráficos no centro), enquanto os modos estáticos permanecem inalterados (gráficos topo).
O modo dinâmico z c é apenas ligeiramente menor que y c (curvas azuis) nos gráficos centrais 5A e 5B . Isso contrasta com os perfis simétricos e imutáveis de ambos (yc , z c ) modos dinâmicos para espermatozóides detectados longe do vidro (curvas vermelhas nos gráficos no centro 5A e 5B ).
Se prestarmos atenção ao sistema de referência de rolamento que o acompanha, a natureza do efeito do vidro é bastante anisotrópica, uma vez que afeta apenas um plano de rolamento, ou seja, no plano b ( 5C ).
A lamínula afeta os modos y cr estático e dinâmico (gráfico acima e no centro a 5C ). Mas o plano z (z cr ) permanece inalterado (gráfico no topo e no centro em 5D ).
A forma do modo estático no sistema de referência de rolagem complementar é a mesma entre todos os espermatozóides e define uma espiral destra fora do centro indicada por h (s) e mostrada pela curva preta em 5E . A projeção da espiral no plano de rolamento (plano verde) é uma espiral no sentido anti-horário que não tem simetria polar, ou seja, deslocado para um lado (projeção cinza em 5E ).
As espirais de modo estático se parecem muito com as espirais logarítmicas que são freqüentemente encontradas na natureza. No entanto, neste caso, o raio da espiral muda de forma não monotônica, aumentando / diminuindo em uma taxa mais rápida do que as espirais logarítmicas em outras partes da natureza.
Espiral logarítmica direita h (s) em 5Epode ser expresso em termos de seu raio e passo ( 5G ), que decai exponencialmente ao longo do comprimento do flagelo. Qualquer mudança no sinal em h y ou h z faz com que a espiral mude para a rotação no sentido horário, criando assim uma espiral para a esquerda. Todos os espermatozóides flutuantes livres geraram espirais no sentido anti-horário. Apenas dois espermatozóides apresentaram espirais no sentido horário, embora com formato idêntico ao 5E . Foram essas duas células que não conseguiram nadar para frente devido a alguns obstáculos em seu caminho, mas podiam continuar a girar em torno de seu próprio eixo de torção.
Em todos os casos, a rotação da hélice se correlaciona com a direção do rolamento do esperma da seguinte maneira: uma hélice no sentido anti-horário para rotação no sentido horário (como visto da extremidade traseira) e uma hélice no sentido horário para rotação no sentido anti-horário.
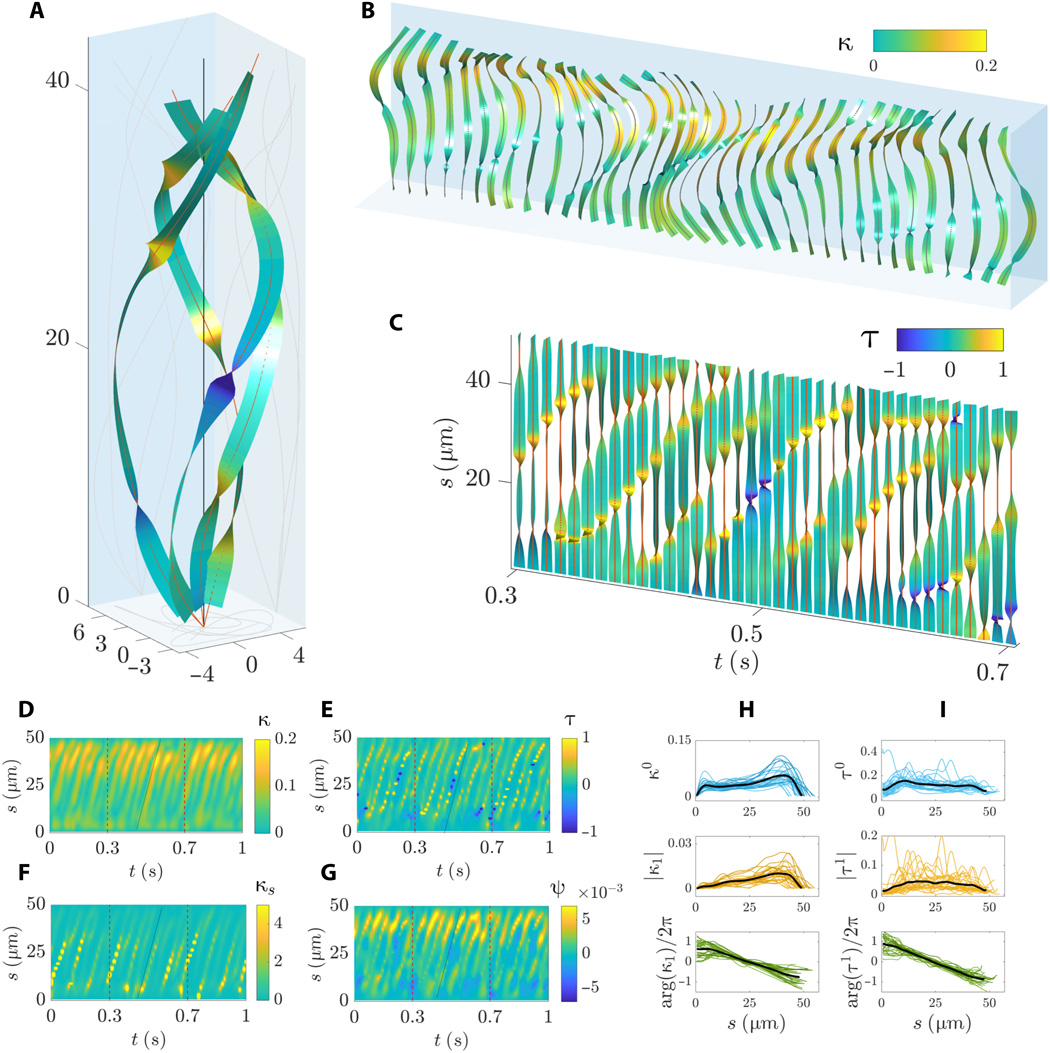
Imagem # 6 Os
gráficos 6A e 6B mostram uma sequência complexa de ondas progressivas conforme o flagelo gira em torno de seu eixo de rolamento. As ondas flexurais se propagam linearmente ao longo do flagelo com uma amplitude não monotônica ao longo do comprimento do arco, caracterizada por um aumento acentuado nas regiões média e distal.
A torção da forma de onda é caracterizada por curvas acentuadas ao longo do comprimento do arco ( 6B e 6C) com voltas positivas e negativas simultâneas. A forma espiral da linha central do flagelo sofre um fenômeno pelo qual coexistem regiões de quiralidade oposta ao longo do flagelo. No entanto, porções do flagelo com quiralidade oposta se movem durante o flapping ( 6C e 6E ). As ondas de torção que se propagam se propagam à mesma velocidade que a onda de curvatura.
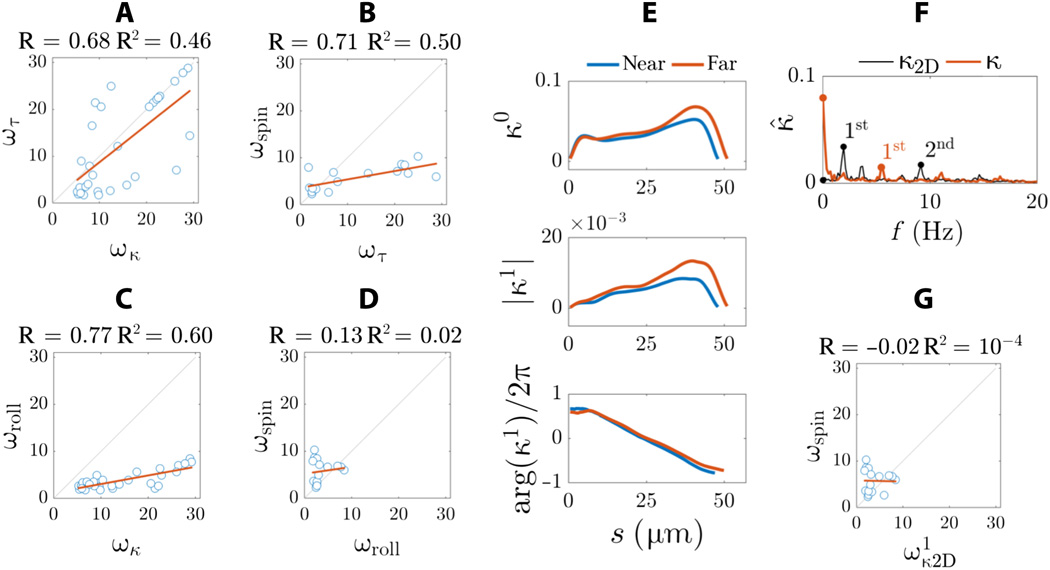
Imagem # 7
Para entender melhor a diferença entre os resultados da microscopia 3D obtidos neste estudo e os resultados da microscopia 2D que são frequentemente descritos na literatura, os cientistas criaram uma projeção 2D a partir de uma forma de onda 3D.
O regime estático da curvatura bidimensional é muito insignificante ( 7F) Em 2D, a assimetria interna da forma de onda não pode ser detectada. Em vez disso, o espectro de frequência é caracterizado por dois picos de frequência (marcadores pretos em 7F ) em vez de um pico de frequência principal visto para curvatura tridimensional (curva vermelha em 7F ).
Para um conhecimento mais detalhado das nuances do estudo, recomendo que você leia o relatório dos cientistas e materiais adicionais a ele.
Epílogo
Nesse trabalho, os cientistas conseguiram mostrar na prática que devido à microscopia tridimensional é possível descobrir o que a microscopia bidimensional não dá - por exemplo, ver mudanças na direção da espiral (mudanças na quiralidade) durante a oscilação dos flagelos.
À primeira vista, pode parecer que os espermatozóides estão simplesmente flutuando para a frente, abanando o rabo. No entanto, um exame detalhado da cinemática dessas células mostrou que esse processo é muito mais complicado. O bater e balançar dos flagelos, a rotação da própria célula - tudo isso junto permite que o esperma avance.
Algumas dessas informações estavam disponíveis anteriormente, mas nem todos os detalhes foram detalhados devido às limitações da microscopia 2D. Entender como os espermatozoides se movem pode ajudar muito no campo da medicina reprodutiva, dizem os cientistas. Em particular, os novos dados irão expandir a gama de parâmetros pelos quais as células sexuais não saudáveis dos homens são determinadas, que é a causa da infertilidade em cerca de metade dos casais.
Em todo caso, quanto melhor compreendermos certos fenômenos e processos, especialmente aqueles que ocorrem em nosso próprio corpo, mais provável será que melhore a qualidade de nossa vida.
Obrigado pela atenção, fiquem curiosos e tenham uma boa semana de trabalho, pessoal. :)
Um pouco de publicidade
Obrigado por ficar com a gente. Você gosta de nossos artigos? Quer ver um conteúdo mais interessante? Ajude-nos fazendo um pedido ou recomendando a amigos VPS em nuvem para desenvolvedores a partir de US $ 4,99 , um análogo exclusivo de servidores básicos que inventamos para você: The Whole Truth About VPS (KVM) E5-2697 v3 (6 núcleos) 10 GB DDR4 480 GB SSD 1 Gbps de $ 19 ou como dividir o servidor corretamente? (opções disponíveis com RAID1 e RAID10, até 24 núcleos e até 40 GB DDR4).
O Dell R730xd 2x é mais barato no data center Equinix Tier IV em Amsterdã? Apenas temos 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 TV a partir de US $ 199 na Holanda!Dell R420 - 2x E5-2430 2.2 Ghz 6C 128 GB DDR3 2x960 GB SSD 1 Gbps 100 TB - a partir de $ 99! Leia sobre como construir a infraestrutura do prédio. classe com servidores Dell R730xd E5-2650 v4 a um custo de 9.000 euros por um centavo?